重磅!中国大陆首个PD-1抑制剂开卖!深度解密其研发细节
28日,中国大陆首个PD-1抑制剂(nivolumab)正式开卖,非小细胞肺癌患者终于用上了期待已久的“O”药。
这是个历史性的时刻。

一周前,百时美施贵宝(BMS)公布了nivolumab在中国大陆的销售价格——只有美国的一半,也远远比其他国家和地区代购的便宜。这个价格获得了满堂彩。
两个月前,nivolumab获中国国家药品监督管理局(CNDA)批准,成为第一个登陆中国大陆的免疫检查点抑制剂。
三年前,在广东省人民医院终身主任吴一龙教授的主持下,CheckMate-078研究开始了。作为nivolumab的中国注册临床研究,它是第一个,也是迄今为止唯一一个证实免疫检查点抑制剂在中国人群中疗效和安全性的研究[1]。CheckMate-078开启了专属于中国晚期非小细胞肺癌患者的生命奇迹。
传奇一般的“O”药究竟是如何诞生的呢?
曙光
癌症,说到底就是人体免疫系统与异常细胞之间的一场战斗。癌症会不会发生,看的就是在这场战斗里谁强谁弱。
一百多年来,科学家一直在探索激活人体免疫系统以治疗癌症的免疫疗法,希望初现于1996年。
当年,加州大学伯克利分校的James Allison团队在《科学》杂志上发表了一篇开天辟地的研究论文,他们在T细胞上发现了一种抑制分子CTLA-4,阻断这种分子与抗原呈递细胞上相应配体分子的结合,就可以增加T细胞的抗肿瘤活性,抑制肿瘤生长[2]。
肿瘤免疫治疗呼之欲出。
就在此时,一直埋头做转基因小鼠技术研发的公司Medarex(2009年被BMS收购)嗅到癌症治疗要巨变的气息。没想到他们不仅嗅觉灵敏,动作还很迅速,很快就开发出了CTLA-4抗体。2000年,他们在转移性黑色素瘤患者身上,亲眼见证了免疫治疗的威力。这个抗体就是2011年被FDA批准的全球第一个免疫检查点抑制剂Yervoy(ipilimumab)。
ipilimumab的成功,让Medarex的科研人员意识到,CTLA-4和类似的靶点必将是肿瘤治疗十分有潜力的新方向。
在Medarex转基因小鼠技术带头人Nils Lonberg的带领下,一帮科研人员开始在浩瀚的科研论文中搜寻新线索。很快,他们就被日本京都大学Tasuku Honjo团队在1992年克隆到的PD-1分子吸引住了[3]。
华人免疫学家陈列平和Tasuku Honjo的研究都表明,这个PD-1分子一旦与肿瘤细胞上的PD-L1结合,T细胞的抗肿瘤活性就没了[4-6]。这就意味着,PD-1和CTLA-4类似,也是T细胞的一种负调节因子。
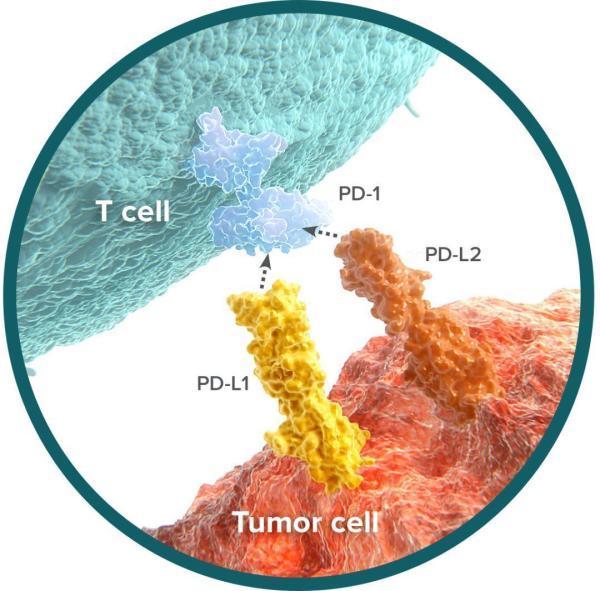
PD-1/PD-L1通路抑制T细胞活性的机制示意图
如果CTLA-4抗体能激活T细胞的抗肿瘤活性,那么PD-1抗体应该也能达到同样的效果!这让Nils Lonberg团队欣喜若狂。
而PD-1抗体开发的任务,就落到了当时在Medarex任职的华人科学家王常玉和黄海春等人的肩上。
全人源才更有效安全
既然要研发可以在人身上使用的PD-1抗体药物,那根据抗原抗体反应的原则,第一步就是获取抗原,即人的PD-1。这个很简单,毕竟早在1992年Tasuku Honjo就已经克隆到了。
接下来问题就来了,抗体是抗原刺激免疫系统之后,体内B细胞根据抗原的特点定制化生产的,抗原PD-1是有了,那该用PD-1刺激谁产生抗体呢?
刺激人?这个你想都不要想。
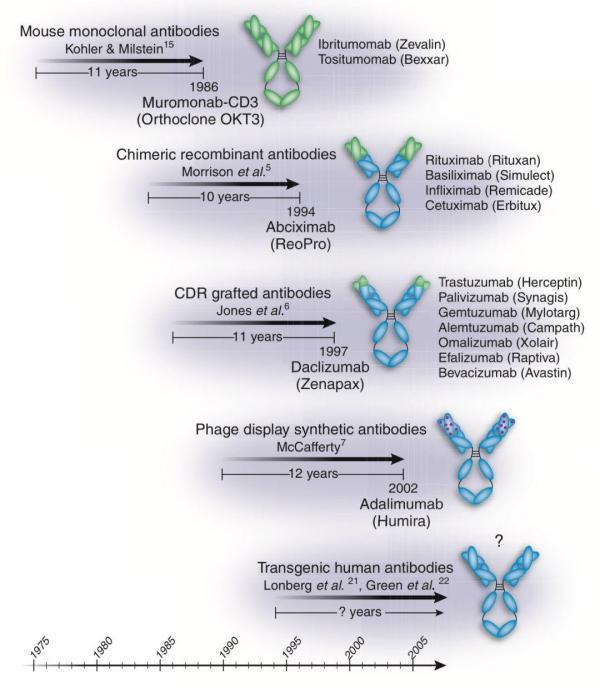
那只能是小白鼠了。这个生产抗体的办法是Kohler和Milstein在1975年首次发现的[7],1984年他们因为发现了单克隆抗体产生原理而获得了诺贝尔生理学或医学奖[8]。

但是这里面就有个问题了。用人的PD-1去刺激小白鼠生产PD-1抗体,这个特异性的结合应该是没问题的。但是呢,当我们把小鼠产生的鼠源PD-1抗体用在人身上的时候,这个鼠源PD-1抗体显然不是人身上的东西啊,人体肯定会把鼠源PD-1抗体当做抗原清除掉[9-11]。可能不起效不说,这种抗体还会带来一些安全问题[12]。
如此一来,这类抗体实用性到底怎么样就说不好了,1986年获FDA批准的第一个单克隆抗体药物Muromonab-CD3就是个明证。它获批之后,FDA连续8年没再敢批其他的单抗药物。
为了尽可能避免人体把鼠源抗体当做外来的抗原清除掉,科学家着实想了很多办法。
1984年,人-鼠嵌合抗体诞生了[13],科学家把鼠源抗体专门用来识别抗原的可变区保留,把对抗原识别作用不大的恒定区用人的抗体替换掉。如此一来,这个人-鼠嵌合抗体进入人体后就没那么招摇了,在一定程度上降低了被人体消灭掉的风险。
抗体药物发展简史
两年后的1986年,Jones等人对鼠源抗体的可变区做进一步改造,让它在保留识别抗原能力的前提下,尽可能的小。这就是我们现在常说的人源化抗体。
不过,就算做了这么多改造,但是显而易见的是,如果能让小鼠生产人的抗体,那才是真的完美。
早在1985年,Frederick Alt等就曾预言,日渐成熟的转基因技术可以让小鼠表达人的抗体[14]。可以说,Alt的预言不仅给了科学家信心,还为未来的研究指明了方向。
1994年,Alt的预言实现了。4月28,当年尚在GenPharm公司(1997年被Medarex收购)工作的Nils Lonberg等人[15],和Cell Genesys公司的研究团队[16],分别在《自然》和《自然遗传学》上发表重要研究成果:他们用不同的方法开发出了可以生产全人源单克隆抗体的转基因小鼠。
各种类型抗体的结构
从Alt的预言,到技术实现,消耗了无数科学家的近十年光阴,治疗性抗体的生产终于近乎完美了。无怪乎有人称:转基因小鼠技术的实现是单克隆抗体制备领域的一次技术飞跃。
制造PD-1抗体,万事俱备,只欠东风。
nivolumab诞生记:精益求精的单抗技术
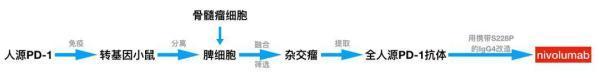
王常玉和黄海春等人抓了几只Nils Lonberg研发的转基因小鼠,把PD-1注射到转基因小鼠体内。那是不是就可以坐等从转基因小鼠体内收获全人源的PD-1抗体了呢?
当然不是。
一只小鼠才能生产多少抗体?此外,用小鼠生产,这成本肯定得高得吓人啊。
研究人员把接受PD-1免疫后的转基因小鼠的脾脏取出来了。很多产生全人源PD-1抗体的B细胞都在里面。让这些脾细胞生产抗体虽然是个好主意,然而这些离体细胞的寿命不长、增殖能力也很差。
要说长寿且繁殖力强,那肿瘤细胞肯定冠绝细胞界。
科学家就想到利用癌细胞的这一特点。他们把脾细胞和骨髓瘤细胞融合,让这种融合细胞既能产生全人源PD-1抗体,还能“健康长寿”。这就是大名鼎鼎的“杂交瘤”技术。通过简单的筛选,王常玉和黄海春团队很快得到了合适的杂交瘤细胞,终于能够生产梦寐以求的全人源PD-1抗体。
此时,距离他们开始制备PD-1抗原只过了4个月!
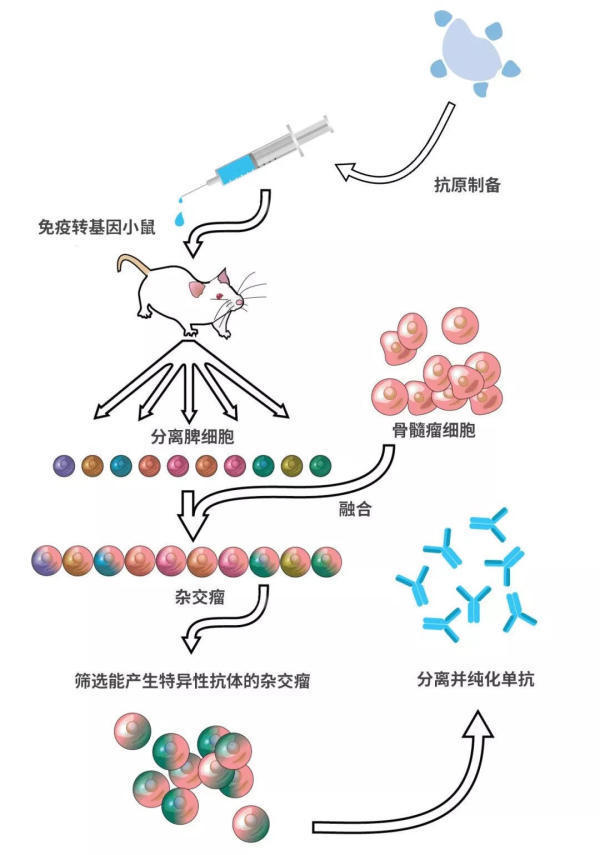
利用转基因小鼠制备抗体的流程图(图:wiki)
不过此时,这个全人源PD-1抗体还不是真正的nivolumab,只能算个“原型机”,要能够稳定、有效地在临床上使用,还得给它动几个“大手术”才行。
这里我们就很有必要先来了解一下抗体本身了。
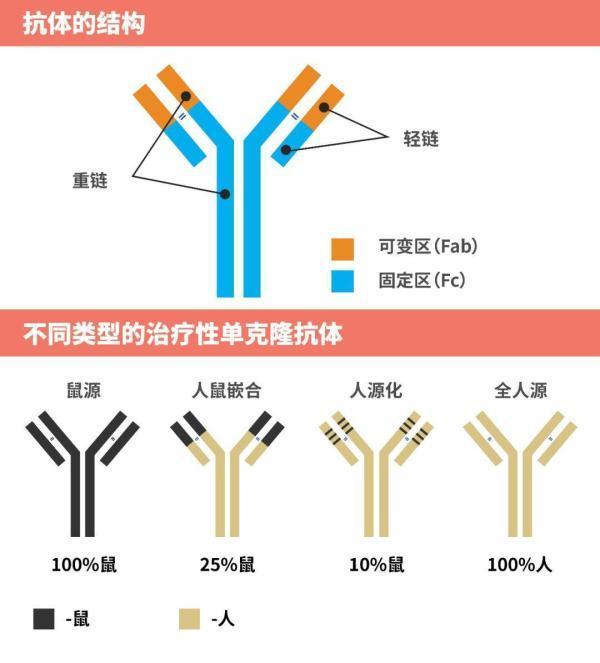
我们都知道抗体和抗原的结合是非常有特异性的,靠着抗体独一无二的可变区,大多数情况下一种抗体能够专一识别一种抗原。正是这种一一对应的关系,让抗体药物有更小的副作用和更高的效率。
不过从另一个层面来分,人体内的抗体又可以分为IgA、IgD、IgE、IgG 和 IgM 等 5 种类型。这五种类型的抗体各司其职,针对不同的外来入侵物,它们的寿命(半衰期)也不同,其中IgG的寿命最长,所以目前临床上使用的治疗性抗体多为 IgG 类型[17]。
这个IgG门下又分四大金刚,它们分别叫IgG1、IgG2、IgG3 和 IgG4。这四种亚型的技能也是不同的[17]。
例如IgG1和IgG3,它俩有两个独门技能,叫抗体依赖的细胞毒性(ADCC)以及补体依赖的细胞毒性(CDC),这两个技能那可是非常厉害啊,一旦IgG1和IgG3与目标细胞上的抗原结合,这个细胞肯定会“中毒”身亡。显然,这种类型的抗体不适合作为T细胞表面抗原PD-1的抗体。毕竟我们的目的是为了激活T细胞,而不是干掉T细胞。
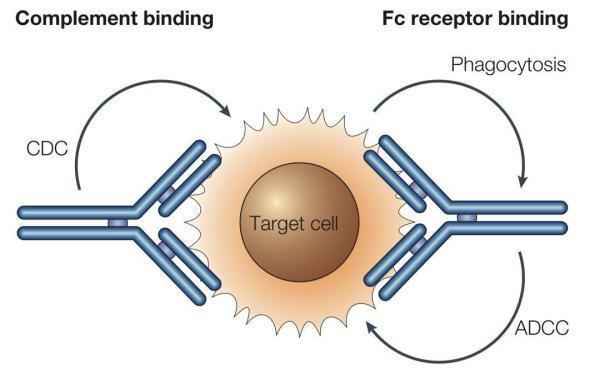
IgG1和IgG3的恒定区(FC)有抗体依赖的细胞毒性(ADCC)何补体依赖的细胞毒性(CDC)
而IgG2和IgG4亚型就温和多了,它俩主要的技能是阻碍或抑制。巧了,科学家不正是要阻碍肿瘤细胞的PD-L1与T细胞的PD-1结合嘛!就是它俩了。不过,IgG2主要结合糖类抗原,所以它也出局了。
如此一来,IgG4亚型的PD-1抗体就是科学家最想要的。
但问题也随之而来了,IgG4亚型的PD-1抗体不是你想要就能要的啊。我们把PD-1抗原注射到小鼠体内,连小鼠自己都不清楚自己会产生哪种抗体,我们就更没办法知道了。
王常玉和黄海春想了个好办法。
既然抗体识别抗原主要依赖于可变区,那把筛选到的那个PD-1抗体的可变区安装到人的IgG4型抗体上,不就行了嘛!研究人员就给全人源PD-1抗体测了个序,确定这个抗体的可变区序列,然后把这个序列和IgG4亚型抗体的恒定区序列连在一起,一个全新的IgG4型的全人源PD-1抗体就可以诞生了。
抗体示意图
好,现在我们距离nivolumab已经很近了!
为啥只是很近了呢,因为王常玉和黄海春还注意到一个问题,那就是野生型(人体正常情况下的)IgG4的两个识别抗原的可变区会发生交换,形成单价(半抗体分子)或者进一步形成双特异性抗体[18]。
一旦上面的那个IgG4型的PD-1抗体与体内的IgG4型其他抗体相遇,它们之间会交换可变区,这显然不利于抗体药物的稳定性,会降低治疗效果;此外,可变区交换后的IgG4抗体,它结合的还是不是PD-1就不好说了,所以这种可变区的交换还会导致不良反应[19]。
对一款药物来说,这是不可接受的。
好在这个问题有解决的办法,科学家早就发现,只要把IgG4恒定区的第228位氨基酸从丝氨酸(Ser)换成脯氨酸(Pro),就可以阻止IgG4抗体之间的可变区交换了。
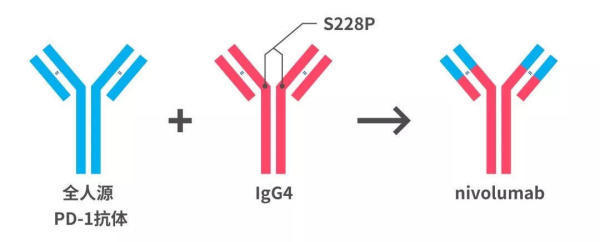
nivolumab的诞生
经过“换头”“整容”的重组抗体——目前唯一的全人源、IgG4型的PD-1单克隆抗体药物nivolumab终于诞生了。
征战临床:千锤百炼的临床证据
在从实验室走向市场之前,更多的考验等待着初生的nivolumab [20]。
对一个好抗体来说,首要的是安全,这个抗体不能危害到其他正常的细胞组织和器官。于是研究人员把包括小脑、心脏、肝、肺、肾和脾等在内的人体细胞和nivolumab放一起。很好,即使在高于使用浓度数十倍水平的情况下,nivolumab也没有伤害上述的组织细胞。
其次,就是抗体的有效性,看看nivolumab是不是真的可以激活T细胞的抗肿瘤能力吧。当把T细胞和一群抑制它活性的细胞一起培养,再往里面加入nivolumab后,T细胞释放γ-干扰素和IL-2的能力显著增强,T细胞的增殖能力增强。这说明T细胞活性增强了。
而且Nivolumab亲和力非常强,可以在非常低的浓度下(~1.5 ng/mL)解除PD-L1对T细胞的抑制,增强T细胞的活性。与此同时,在没有抗原或者T细胞受体存在的情况下,nivolumab不影响T细胞的活性。
和预期的一样,在0.003到50μg/mL浓度区间内,nivolumab对T细胞没有出现ADCC和CDC,它确实不会伤害T细胞。
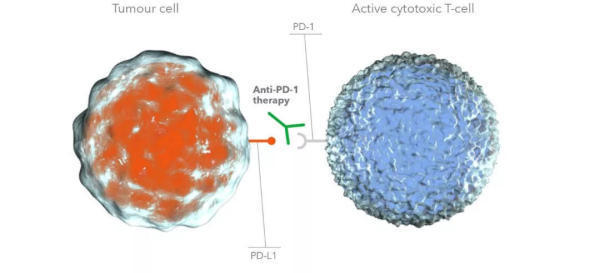
PD-1抗体阻断PD-L1与PD-1的结合
这些结果表明nivolumab不会引起非特异性的淋巴细胞活化。
可见nivolumab不仅高效,而且效果特异性高,只管给肿瘤特异性T细胞加油打气从不做小动作。至此所有的临床前研究都准备好了,就等上临床来个终极一战。
2006年,在王常玉和陈列平等科学家和医生的推动下,全球第一个PD-1抗体临床试验启动了[21]。这个研究确定了nivolumab在人体的安全性,以及有效性。
2008年开始nivolumab里程碑式的CA209-003研究,确定了nivolumab的临床使用剂量为3mg/kg [24]。同时,CA209-003研究证实nivolumab在多个瘤种中具有抗肿瘤作用,全面开启临床之路,截止目前累计发表15篇NEJM文章。
2014年7月4日,nivolumab率先在日本获批,成为全球第一个获批的PD-1抑制剂。自此,nivolumab凭借出生自带的光环和荣耀,在抗肿瘤征途上一路开挂。
短短4年内,nivolumab在全球已获批用于9个瘤种,17项适应症,是至今全球适应症最多的PD-1抑制剂。

从呱呱坠地,到为抗击肿瘤一路征战,nivolumab自其诞生之日起就自带“光环”,她的传奇之旅仍将继续。
参考资料:
[2]. Leach D R, Krummel M F, Allison J P, et al.Enhancement of Antitumor Immunity by CTLA-4 Blockade.[J]. Science, 1996, 271(5256): 1734-1736.
[3]. Ishida Y, Agata Y, et al. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death.[J]. The EMBO Journal, 1992, 11(11): 3887-3895.
[4]. Nishimura H, Nose M, Hiai H, et al. Development of Lupus-like Autoimmune Diseases by Disruption of the PD-1 Gene Encoding an ITIM Motif-Carrying Immunoreceptor[J]. Immunity, 1999, 11(2): 141-151.
[5]. Dong H, Zhu G, Tamada K, et al. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion.[J]. Nature Medicine, 1999, 5(12): 1365-1369.
[6]. Freeman G J, Long A J, Iwai Y, et al. Engagement of the Pd-1 Immunoinhibitory Receptor by a Novel B7 Family Member Leads to Negative Regulation of Lymphocyte Activation[J]. Journal of Experimental Medicine, 2000, 192(7): 1027-1034.
[7]. Kohler G, Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity[J]. Nature, 1975, 256(5517): 495-497.
[8].The Nobel Prize in Physiology or Medicine 1984
[9]. Pendley C, Schantz A, Wagner C, et al. Immunogenicity of therapeutic monoclonal antibodies.[J]. Current Opinion in Molecular Therapeutics, 2003, 5(2).
[10]. Ortho Multicenter Transplant Study Group. A randomized clinical trial of OKT3 monoclonal antibody for acute rejection of cadaveric renal transplants[J]. New England Journal of Medicine, 1985, 313(6): 337-342
[11]. Kuus-Reichel K, Grauer L S, Karavodin L M, et al. Will immunogenicity limit the use, efficacy, and future development of therapeutic monoclonal antibodies?[J]. Clinical and diagnostic laboratory immunology, 1994, 1(4): 365-372.
[12]. Baert F, Noman M, Vermeire S, et al. Influence of immunogenicity on the long-term efficacy of infliximab in Crohn's disease[J]. New England Journal of Medicine, 2003, 348(7): 601-608.
[13]. Boulianne G L, Hozumi N, Shulman M J, et al. Production of functional chimaeric mouse/human antibody[J]. Nature, 1984, 312(5995): 643-646.[].
[14]. Alt F W, Blackwell T K, Yancopoulos G D, et al. Immunoglobulin genes in transgenic mice[J]. Trends in Genetics, 1985: 231-236.
[15]. Lonberg N, Taylor L D, Harding F A, et al. Antigen-specific human antibodies from mice comprising four distinct genetic modifications.[J]. Nature, 1994, 368(6474): 856-859.
[16]. Green L, Hardy M C, Maynardcurrie C E, et al. Antigen–specific human monoclonal antibodies from mice engineered with human Ig heavy and light chain YACs[J]. Nature Genetics, 1994, 7(1): 13-21.
[17]. Salfeld J G. Isotype selection in antibody engineering[J]. Nature Biotechnology, 2007, 25(12): 1369-1372.
[18]. Davies A M, Rispens T, Heer P O, et al. Structural determinants of unique properties of human IgG4-Fc.[J]. Journal of Molecular Biology, 2014, 426(3): 630-644.
[19]. Labrijn A F, Buijsse A O, Den Bremer E T, et al. Therapeutic IgG4 antibodies engage in Fab-arm exchange with endogenous human IgG4 in vivo[J]. Nature Biotechnology, 2009, 27(8): 767-771.
[20]. Wang C, Thudium K B, Han M M, et al. In Vitro Characterization of the Anti-PD-1 Antibody Nivolumab, BMS-936558, and In Vivo Toxicology in Non-Human Primates[J]. Cancer immunology research, 2014, 2(9): 846-856.
[21]. Brahmer J R, Drake C G, Wollner I, et al. Phase I Study of Single-Agent Anti–Programmed Death-1 (MDX-1106) in Refractory Solid Tumors: Safety, Clinical Activity, Pharmacodynamics, and Immunologic Correlates[J]. Journal of Clinical Oncology, 2010, 28(19): 3167-3175.

来源:奇点网
版权及免责声明:本网站所有文章除标明原创外,均来自网络。登载本文的目的为传播行业信息,内容仅供参考,如有侵权请联系答魔删除。文章版权归原作者及原出处所有。本网拥有对此声明的最终解释权。
{replyUser1} 回复 {replyUser2}:{content}