《Gut》论文深度解读:CPA1突变如何引发慢性胰腺炎
慢性胰腺炎(chronic pancreatitis)是各种病因引起胰腺组织和功能发生不可逆改变的慢性炎症性疾病。它的发作通常是由与消化蛋白酶或其抑制剂相关的遗传因素决定的。PRSS1、SPINK1和CTRC等基因的突变会改变胰蛋白酶水平和胰腺炎风险。
重要的是,慢性胰腺炎的遗传风险也可以通过与胰蛋白酶活性无关的机制来介导,如一些PRSS1突变对胰蛋白酶活性没有影响,但会导致酶错误折叠,引发内质网(ER)应激。同样地,CPA1(羧肽酶A1)突变也在体外引起酶错误折叠,导致CPA1细胞内滞留和降解以及相关的ER应激。
尽管体外观察到的结果支持突变诱导的错误折叠和ER应激,但一直缺乏适当动物模型的证据,而CPA1突变的致病机制也是假设的。这限制了研究人员对疾病机制的了解,也阻碍了新型疗法的临床前试验。
为此,美国波士顿大学的研究人员建立了一个新型的小鼠基因敲入品系,在小鼠Cpa1基因座上携带人CPA1突变p.N256K。通过对这种小鼠品系的研究,他们证实,CPA1突变导致酶错误折叠,并通过ER应激的机制引起慢性胰腺炎。这项重要的成果发表在消化道领域的顶尖杂志《Gut》上,影响因子达17.016。
这项研究的重要意义在于,这是第一个与消化酶错误折叠和ER应激相关的慢性胰腺炎小鼠模型。它特别适合测试各种环境因素(如酒精、吸烟、高脂饮食)或药物对慢性胰腺炎发生和发展的影响。
本文也提供了一个研究新思路,对于一些体外实验已有结论但还缺乏活体实验证实的基因变异疾病,通过构建携带变异的小鼠模型来进行活体研究不失为一种可行的方法。
在这项研究中,波士顿大学的Miklós Sahin-Tóth和Eszter Hegyi委托赛业生物(Cyagen)通过C57BL/6胚胎干细胞的同源重组将人CPA1突变p.N256K敲入小鼠的Cpa1基因座,制备出CPA1点突变小鼠(CPA1 N256K品系)。同时,他们还制备出CPA1 null品系,因为新霉素(neo)表达框的存在导致Cpa1 mRNA和CPA1蛋白表达水平降低。此品系可作为对照,以区分CPA1蛋白活性丧失或CPA1错误折叠。
为了鉴定CPA1 N256K和CPA1 null小鼠品系胰腺中Cpa1 mRNA的表达,研究人员从1月龄的动物中制备胰腺cDNA。定量PCR的分析结果表明,与C57BL/6N对照相比,CPA1 N256K品系的Cpa1 mRNA表达降低了30%,而CPA1 null品系则降低了95%。Western blot分析也表明,CPA1 N256K品系的CPA1蛋白含量虽降低但仍然明显,而CPA1 null品系则检测不到CPA1蛋白的表达(图1)。
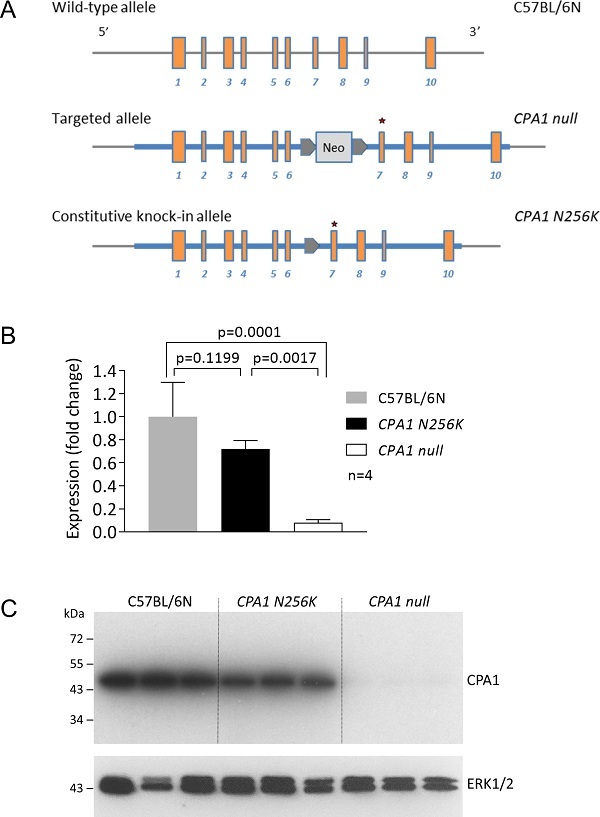
图1. CPA1 N256K小鼠的制备和突变等位基因的表达。(A) 通过同源重组方法制备出CPA1 null和CPA1 N256K品系。(B) 通过定量PCR检测1月龄的CPA1 N256K、CPA1 null和C57BL/6N对照小鼠胰腺中Cpa1 mRNA的表达。(C) 通过western blot分析CPA1 N256K、CPA1 null和C57BL/6N对照小鼠胰腺中的CPA1蛋白表达。
之后,他们从6周龄的CPA1 N256K和C57BL/6N对照小鼠中分离胰腺腺泡,并测定CPA1向培养基中的分泌。与野生型的CPA1蛋白相比,CPA1 p.N256K突变体的分泌几乎检测不到。相比之下,腺泡细胞裂解液的western blot分析显示了高水平的CPA1突变蛋白,表明突变蛋白滞留在细胞内(图2)。同时,他们也注意到N256K品系的细胞内CPA1蛋白水平略低于对照,表明一些错误折叠和滞留的突变蛋白被降解。

图2. CPA1 N256K和C57BL/6N对照小鼠的腺泡细胞分泌的CPA1蛋白(自发或雨蛙素诱导)。(A) 添加或未添加30 pM雨蛙素孵育60分钟后,分泌到培养基中的CPA1。(B) 添加或未添加30 pM雨蛙素孵育60分钟后,细胞裂解液中的CPA1含量。
研究人员发现,两种新的CPA1品系都没有表现出明显的表型改变,而且正常繁殖。CPA1 null小鼠和C57BL/6N对照小鼠的胰腺形态相似。相比之下,CPA1 N256K小鼠从3月龄起就被观察到胰腺偏小。重量测定证实了CPA1 N256K品系的胰腺出现萎缩。为了鉴定胰腺萎缩的组织学变化,他们对1、3、6和12月龄的小鼠胰腺切片进行染色,并将它们与对应的CPA1 null 和C57BL/6N切片进行比较。
对于CPA1 N256K突变小鼠,研究人员观察到慢性胰腺炎的标志,包括腺泡细胞萎缩和炎性细胞浸润。Masson三色染色法进一步证实了这一结论,它揭示了CPA1 N256K小鼠腺泡中的广泛纤维化。通过细胞角蛋白19和Sox9的免疫组化染色,他们还观察到腺泡-导管化生(假管复合物)。相比之下,CPA1 null对照组未表现出胰腺疾病的迹象,与C57BL/6N对照小鼠类似(图3)。
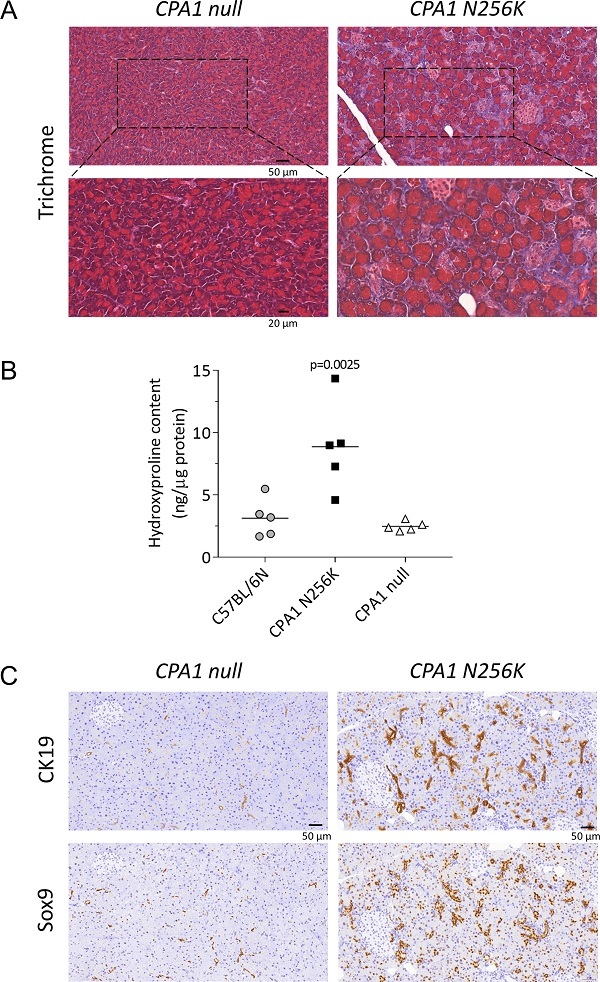
图3. CPA1 N256K小鼠的纤维化和腺泡-导管化生。(A) Masson三色染色法显示CPA1 N256K小鼠的腺泡周围存在广泛的纤维化(蓝色)。(B) CPA1 N256K小鼠胰腺中的羟脯氨酸含量相对于CPA1 null小鼠和C57BL/6N对照增加。(C) CK19和Sox9的免疫组化结果显示CPA1 N256K小鼠存在广泛的腺泡-导管化生,而CPA1 null小鼠没有。
最后,为了确定CPA1 N256K品系的组织学变化是否与ER应激标志物升高有关,研究人员分别测定了1月龄、3月龄和12月龄小鼠的伴侣蛋白Hspa5(BiP)和转录因子Ddit3(CHOP)。与C57BL/6N和CPA1 null对照相比,CPA1 N256K小鼠的BiP表现出一致但适度的上调,而CHOP在所有时间点表现出明显的上调(图4)。
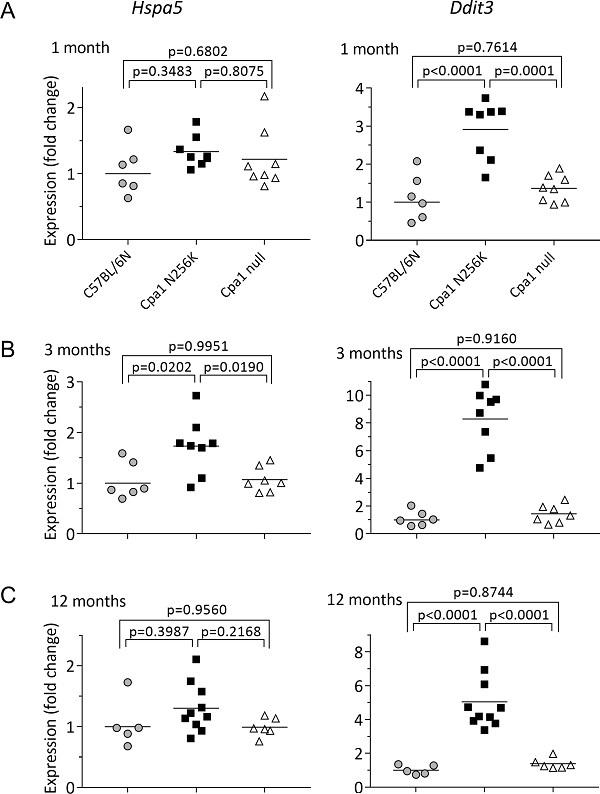
图4. CPA1 N256K小鼠、CPA1 null小鼠和C57BL/6N对照小鼠的内质网应激标志物Hspa5和Ddit3的表达,A、B和C分别对应1月龄、3月龄和12月龄。对胰腺cDNA样本进行定量PCR,结果表示为相对于C57BL / 6N结果平均值的倍数变化。
CPA1 N256K基因敲入小鼠上的一系列实验表明,CPA1突变p.N256K导致酶错误折叠,并引发慢性胰腺炎,同时伴随着ER应激相关的促凋亡转录因子CHOP的上调。CPA1 null小鼠并没有观察到这些病理现象,表明CPA1功能丧失本身并不引起腺泡细胞损伤。错误折叠依赖性的通路在慢性胰腺炎中发挥致病作用,这也为针对ER应激相关机制的治疗干预打下基础。
研究人员强调,这种错误折叠诱导的慢性胰腺炎新模型有望在临床前研究中发挥作用。CPA1 N256K小鼠品系正常发育和繁殖。这些特征以及胰腺病变的时间线使其成为测试各种环境损害(酒精、吸烟、高脂饮食)或药物对慢性胰腺炎影响的理想模型。
相关论文:
Human CPA1 mutation causes digestive enzyme misfolding and chronic pancreatitis in mice
Gut 2018;0:1–12. doi:10.1136/gutjnl-2018-315994

来源:生物通
版权及免责声明:本网站所有文章除标明原创外,均来自网络。登载本文的目的为传播行业信息,内容仅供参考,如有侵权请联系答魔删除。文章版权归原作者及原出处所有。本网拥有对此声明的最终解释权。
{replyUser1} 回复 {replyUser2}:{content}