基因编辑治疗时代已来: 三大基因编辑治疗策略
近日,基因编辑技术迎来了重要里程碑。Verve Therapeutics 公司宣布,其开发的碱基编辑疗法 VERVE-101 在新西兰获批开展人体临床试验,用于治疗杂合子家族性高胆固醇血症(HeFH),这也是首个体内碱基编辑的临床试验。目前已有几种类型的基因编辑剂已被用于体内基因编辑治疗,本文就常用的三大基因编辑治疗策略做一个简单介绍(图1)。
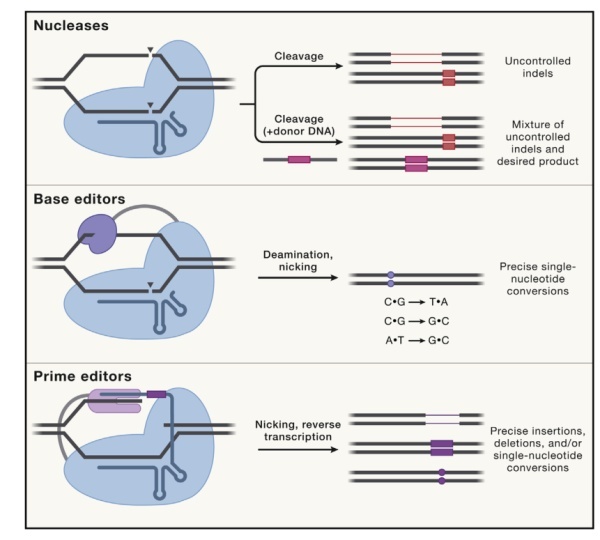
图1. 治疗性基因编辑三大策略
目前,基本上在哺乳动物细胞中进行基因编辑都涉及到使用核酸酶,其在基因组DNA的特定位置产生双链断裂(DSB)。归巢核酸内切酶(Meganuclease)、锌指核酸酶(ZFN)和转录激活物样效应核酸酶(TALEN)是第一批用于哺乳动物细胞基因编辑的酶(图2)。然而,由于这些编辑剂在切割前依赖复杂的蛋白质:DNA相互作用来结合特定的DNA序列,因此必须在新的基因组目标位置设计和构建新的编辑蛋白质。这些方法是非常耗时、耗力的。
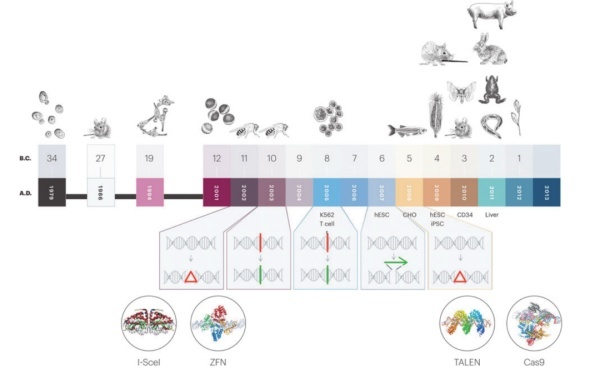
图2. 用于基因编辑的核酸酶发展时间线
在哺乳动物细胞中,DSB最常见的修复方式是非同源末端连接或微同源介导的末端连接,这两种方法都会导致靶点上插入和缺失的不可控分布。在某些情况下,DSB可以通过同源定向修复。这一过程可以由外源DNA供体作为模板,该供体可以包含任何任意序列,两侧是与靶点同源的区域,原则上可以用于在基因组DNA中安装任何所需的序列。然而,同源定向修复在大多数细胞类型中都是低效的,包括非分裂细胞,并且成功的同源定向修复编辑结果通常伴随着大量的插入或缺失改变。出于这些原因,对于将突变的基因精确地纠正为正确的序列,并最大限度地减少不需要的编辑副产品,核酸酶介导的基因编辑通常是一种很差的通用策略。然而,核酸酶通过引入移码突变来扰乱基因的编码序列,或者破坏非编码序列中的调控基序来调节基因的表达是十分有效的。在用核酸酶进行的基因编辑中,目前绝大多数治疗方式是破坏目标基因组位点。
尽管可编程核酸酶已经被应用于治疗,但也有报道称,核酸酶介导的基因编辑会产生一些不良后果。除了靶点的插入或缺失外,在基因组DNA中产生DSB还会导致大的缺失、染色体易位等。这些缺点,促使了更精确的基因编辑替代策略的发展。
碱基编辑
碱基编辑克服了核酸酶的许多限制,通过基因组DNA中的单核苷酸转换实现了精确的基因校正,而不需要DSB。碱基编辑是由融合到可编程DNA结合域的DNA修饰酶组成的。迄今已有许多不同类型的碱基编辑被报道。最早报道的碱基编辑是胞嘧啶碱基编辑分子,它执行靶向的C·G向T·A的转换,由胞苷脱氨酶融合到催化受损的Cas酶和尿嘧啶糖基酶抑制物(UGIs)组成。在典型的胞嘧啶碱基编辑中,催化受损的Cas酶首先与特定的基因组位点结合,而不产生DSB。引导RNA和靶DNA链之间的碱基配对暴露出单链DNA bubble,该bubble可被融合的胞苷脱氨酶结构域进行脱氨基。由于融合的胞苷脱氨酶是单链DNA底物所特有的,脱氨作用被限制在暴露的DNA链内的一个小窗口内。胞嘧啶的脱氨基产生尿嘧啶,它被融合的UGIs部分保护而不被切除,导致目标DNA位点上的U·G错配。催化受损的Cas酶选择性地只切割未编辑的含有G的链而不产生DSB,这会偏向细胞错配修复。由此产生的 U·A 碱基对最终通过细胞修复机制转化为 T·A 碱基对。如果含尿嘧啶的中间体没有受到保护,而是促进了尿嘧啶的切除,结果通常是 C·G向G·C 转化,而不是 C·G向T·A 转化。这一现象被用来设计基于C·G到G·C的编辑程序。
腺嘌呤碱基编辑通过类似的机制执行A·T到G·C转换。由于目前还没有已知的天然酶催化脱氧腺苷进行脱氨基,迄今为止描述的所有腺嘌呤碱基编辑都是使用实验室进化的脱氧腺苷脱氨酶。腺嘌呤碱基编辑是一类特别有用的碱基编辑程序,因为它们逆转了最常见的致病点突变类型(C·G 向T·A),这种突变约占已知致病单核苷酸多态(SNPs)的一半.
自从最初的胞嘧啶碱基编辑和腺嘌呤碱基编辑的开发以来,许多实验室已经报告了数百个不同性质的碱基编辑。碱基编辑已被用于应用各种体外和体内编辑治疗,以纠正致病点突变或或插入可预防或挽救疾病表型的单核苷酸变体。
先导编辑(Prime editors)
虽然碱基编辑原则上可以纠正大多数致病SNPs,但它们不能执行所有可能的单核苷酸转换,也不能介导靶向插入或缺失。为了解决这些限制,研究人员开发了先导编辑器,它允许编程安装任何单核苷酸转换、小的插入、删除或其组合,而不会产生DSB。
先导编辑器由一个融合到Cas9切口酶结构域的逆转录酶组成,并使用一个经过工程化的先导编辑引导 RNA(pegRNA)来将Cas9切口酶定向到特定的靶点,进而对感兴趣的碱基进行编辑。先导编辑器首先切割非靶标DNA链,并使用产生的自由3’端以pegRNA延伸为模板启动逆转录。在将所需的编辑加入到新合成的链之后,可以使用额外的缺口来引导DNA修复,以通过使用编辑的链作为模板来替换未编辑的链。
已经有些研究报道了使用先导编辑器在体内进行编辑的例子。最近对 先导编辑器和 pegRNA 的改进已实现了对各种细胞类型的高效编辑。这将对未来的体内编辑发挥巨大作用。
全世界有数百万人患有遗传疾病,原则上其可以通过治疗性DNA编辑试剂来纠正。CRISPR-Cas技术的广泛发展和优化已经产生了强大的基因编辑工具,包括可编程核酸酶、碱基编辑和先导编辑。将这些基因编辑工具与高效的递送方法相结合已经产生了强大的治疗结果。新的体内基因编辑疗法正迅速走向临床,人类活体基因编辑治疗的时代已经到来。
来源:清风 生物制药小编

来源:网络
版权及免责声明:本网站所有文章除标明原创外,均来自网络。登载本文的目的为传播行业信息,内容仅供参考,如有侵权请联系答魔删除。文章版权归原作者及原出处所有。本网拥有对此声明的最终解释权。
{replyUser1} 回复 {replyUser2}:{content}